
Introduction
Hanwoo (Bos taurus) is a Korean breed of cattle that has been in used as draft animal for 5,000 years (Kim et al., 2011). Hanwoo may have developed independently from an ancestor of the European and African species (Yoon et al., 2005). In Korea, consumer demand is driving efforts to increase meat production and produce higher quality meat (Chung, 2005; Seong et al., 2011). All beef carcasses in Korea are graded by the Korea Institute for Animal Products Quality Evaluation (KAPE; www.ekape.or.kr) both in qualitative and quantative terms. Under KAPE standards (Kim at al., 2011; Choy et al., 2012), the major quantitative components that determine carcass grades are backfat thickness (BF), longissimus dorsi muscle area (LMA), carcass weight (CW) and marbling score (MS).
Earlier reports indicate that single nucleotide polymorphisms (SNPs) in alpha-1-microglobulin/Bikunin precursor (AMBP) gene, a gene in the QTL region (http://www.animalgenome.org/cgi-bin/QTLdb/BT/ link_genom2snpb?chromos=21&upcm=73.81&locm=76.81),are associated with meat quality traits in swine(Cinaretal.,2012). AMBP gene is located on the bovine chromosome 8 and consists of ten exons. It is a single-copy gene and is expressed exclusively in the liver (Grewal et al., 2005; Tyagi et al., 2002), encoding two plasma glycoproteins: alpha-1-microglobulin (A1M) and Bikunin (Grewal et al., 2005; Tyagi et al., 2002; Allhorn et al., 2002). Although A1M and Bikunin originate from AMBP gene cleavage by a furin-like protease, the structural and functional relationship between the two mature molecules remains unknown (Tyagi et al., 2002). The first 6 exons code for A1M, exons 8 and 9 code for each Kunitz domain and exons 7 and 10 encode the N- and C-terminal ends of Bikunin, respectively (Fries and Blom, 2000).
AMBP gene is reported to assumed to be cis-regulated and was part of the glycolysis pathway (Heidt et al., 2013). A1M is to play a role in the regulation of development (Logdberg et al., 2000), cell growth (Logdberg and Akerstrom, 1981; Logdberg et al., 1986), metabolism (Allhorn et al., 2002) and the immune response (Wester et al., 2000; Santin and Cannas, 1999). Bikunin is a urinary trypsin inhibitor belonging to the Kunitz-type superfamily of protease inhibitors (Fries and Blom, 2000). It is thought to participate in the control of such events as endothelial cell growth or extracellular matrix stabilization (Tyagi et al., 2002).
The aim to the study was to detection of novel polymorphisms within AMBP gene by sequencing and to determine the genetic association between the SNPs and economic traits, such meat quality and carcass traits
Materials and methods
Sample collection and Genomic DNA Extraction
A total of 105 Korean cattle (Hanwoo) steers obtained from Korea Institute for Animal Products Quality Evaluation (KAPE) were used as samples. Genomic DNA from tissue samples was extracted using a Toyobo MagExtractor Kit, as described in the manufacturer's instructions (Toyobo, Japan). The DNA was quantified by NanoDrop ND-1000 UV - Vis Spectrophotometer (NanoDrop Technologies, USA). For each specimen, the weight at slaughter was recorded as the CW. BF and LMA were calculated at the 12th and 13th rib interface. MS was estimated on a cross section at the 12th and 13thrib interface and scored on a scale from 1 to 9 associated with the most marbling.
Primer design and Polymerase Chain Reaction
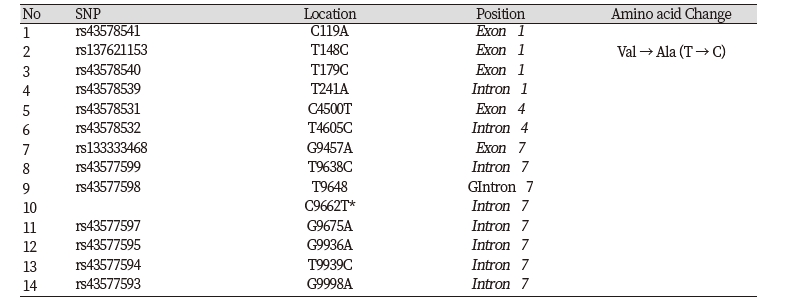
Primers used in the study were based on the nucleotide sequence of the bovine AMBP gene available in the GenBank database (Accession No. AC_000165). Detailed information on oligonucleotide primers, amplicon size and corresponding annealing temperature is depicted in Table 1.

PCR amplification was performed using genomic DNA (20~200 ng/㎕), 10 pmole of each primer, 2 ㎕ of 10X buffer (10 mM Tris-HCl, 50 mM KCl, 1.5 mM MgCl2,pH8.3),40㎛ of dNTPs and 0.5 units Taq polymerase (Genetbio, Korea). Thermal cycling began with an initial denaturation step of 94℃ for 5 min, followed by 35 cycles of 94℃ for 1 min, optimized annealing temperature 63.5℃ for 30 s, 72℃ for 30 s and a final extension at 72℃ for 5 min using a Peltier Thermal Cycler 200 (MJ Research, USA).
Sequencing analysis
The amplified PCR product of each sample was excised from a 2.0% agarose gel and purified with AccuPrep®Gel PurificationKit(Bioneer™, Korea). The purified products were sequenced using BigDye Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, USA). Sequencing reactions were purified by Ethanol and run on ABI 3130 XL Genetic Analyzer (Applied Biosystems, USA) to determine nucleotide sequences. Detection of SNPs were performed with Seq Man II software (DNA Star, USA).
Statistical analysis
Allele and genotype frequencies were calculated by simple counting. A linkage disequilibrium (LD) block of polymorphisms was tested by Haploview version 3.32 (Barrett et al., 2005). The haplotypes and their frequencies were calculated with the expectation-maximization algorithm (Stephens et al., 2001). Statistical analysis of the association between the genotypes (or haplotypes) of A1M and economic traits were evaluated with the least square method (GLM implemented in the SAS software package: SAS institute, USA). The general linear model used in this study was
Yij=μ+Gi+βA+Si+eij,
Where Yijis the observation of the performance traits, μ is the overall mean for each trait, Giis the ith level of the fixed factor genotype, β is the regression coefficient for age, A is the age, Si is the it sex effect and eij is the random error for the ijth individual.
Results
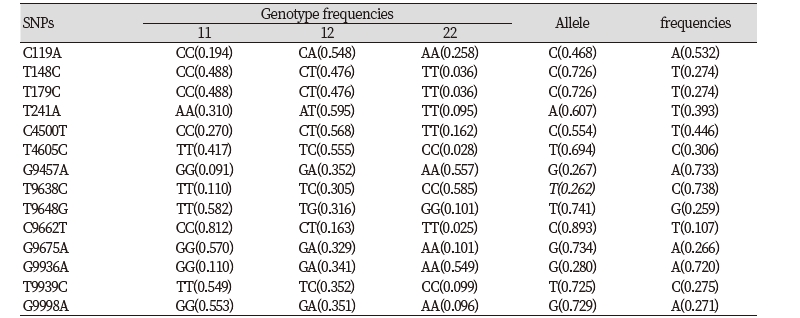
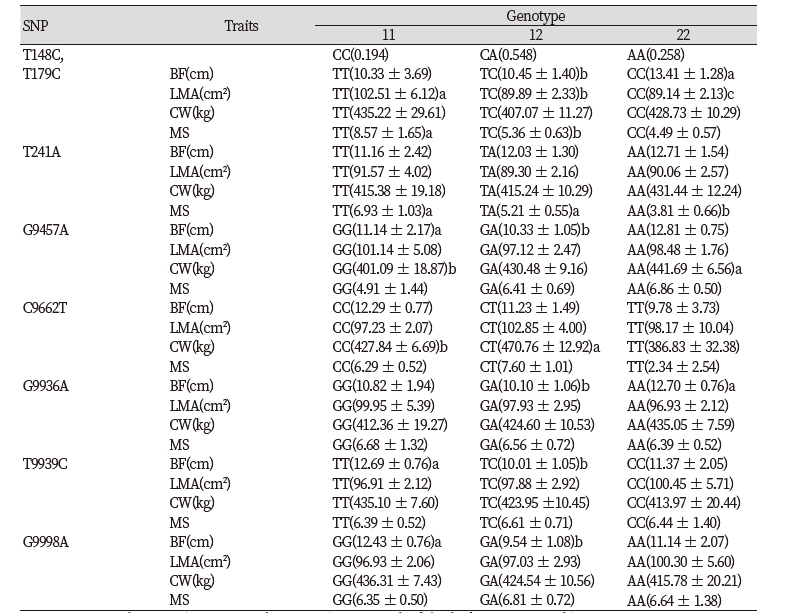
We investigated genetic variations within the bovine AMBP gene using DNA sequencing technologies. By comparing the sequencing results with the DNA sequence of Bos taurus AMBP gene, we identified 14 SNPs. 5 SNPs (C119A, T148C, T179C, G4500T, G9457A) were detected in the exon regions and 9 SNPs (T241A, T4605C, T9638C, T9648G, C9662T, G9675A, G9936A, T9939C, G9998A) were detected in the intron regions (Fig. 1, Table 2). The T148C was missense mutation. T148C changes amino acid from valine to alanine (Table 2). The locations, allele frequency, and genotype frequency for Hanwoo are shown in Table 3. The frequency of T148C and T179C polymorphisms in allele C was higher than that for allele T,and 0.488, 0.476 and 0.036 displayed the CC, CT and TT genotypes, respectively. The frequency of T148C and T179C are the same in genotype. And the genotype frequencies were as follows : 0.194 CC, 0.548 CA and 0.258 AA for the C119A; 0.310 AA, 0.595 AT and 0.095 TT for the T241A; 0.270 CC, 0.568 CT and 0.162 TT for the C4500T; 0.417 TT, 0.555 TC and 0.028 CC for the T4605C; 0.091 GG, 0.352 GA and 0.557AA for the G9457A; 0.110 TT, 0.305 TC and 0.585 CC for the T9638C; 0.582 TT, 0.316 TG and 0.101 GG for the T9648G; 0.812 CC, 0.163 CT and 0.025 TT for the C9662T; 0.570 GG, 0.329 GA and 0.101 AA for the G9675A; 0.110 GG, 0.341 GA and 0.549 AA for the G9936A; 0.549 TT, 0.352 TC and 0.099 CC for the T9939C; 0.553 GG, 0.351 GA and 0.096 AA for the G9998A (Table 3).
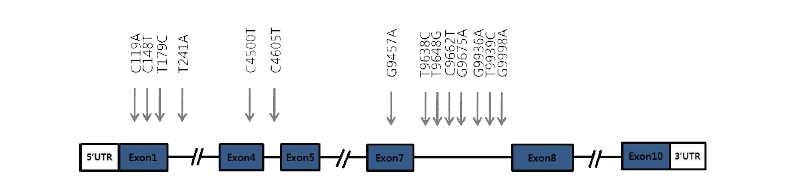
Pairwise LD analysis with the 3 polymorphisms parsed into 1LD block in A1M region and 6 polymorphisms parsed into 2LD blocks in Bikunin region. Three major haplotypes were constructed in A1M region and two major haplotypes were constructed in Bikunin region (frequency < 0.1) (Fig. 2).
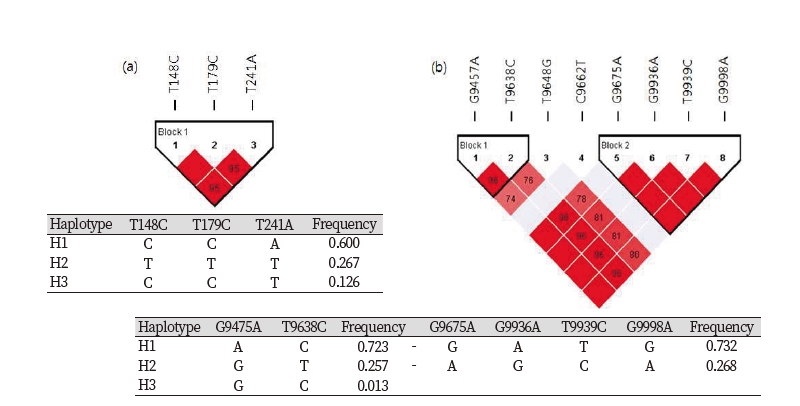
Figure 2. Haplotypes and LD coefficients in AMBP gene. (a) LD blocks comprised 3 polymorphisms of A1M region are shown using Haploview and haplotype frequencies. (b) LD blocks among 8 polymorphisms of Bikunin region are shown using Haploview and haplotype frequencies. The color code on the Haploview plot follows the standard color scheme: shades of red (|D’|< 1, LOD≥2); bright purple (|D'|< 1, LOD< 2).
To investigate the effects of SNPs, we analyzed the association of AMBP genotypes to determine the effects on variations in economic traits. Both the T148C and the T179C SNPs were associated with a significant effect on BF, LMA and MS. Hanwoo with the CC genotype displayed a significantly thicker BF than cattle with genotype TC, Hanwoo with the TT had a larger LMA than those with CC, Hanwoo with TT had a higher MS than those with TC (P< 0.05). The T241A significantly affected MS. Hanwoo with TT and TA had a higher MS than those with AA (P<0.05). The G9457A significantly affected BF and CW. Hanwoo with GG genotype displayed a significantly thicker BF than cattle with GA, Hanwoo with AA had a heavier CW than those with GG (P<0.05). The C9662T significantly affected CW. Hanwoo with CT had heavier CW than those with CC (P<0.05). And G9936A, T9939C, G9998A significantly affected BF (P<0.05) (Table 4).
Discussion
AMBP gene is located on bovine chromosome 8 and SNPs in AMBP gene, a gene in the QTL region, are associated with meat quality traits in swine (Cinar et al., 2012). Therefore, this study aims to investigate the AMBP gene in association to economic traits in Hanwoo.
We show 14 SNPs were detected in the AMBP gene. The T148C displayed a change in amino acid from valine to alanine (Table 2). Based on our statistical association analysis, T148C and T179C were the same in genotype. They also significantly affect BF, LMA and MS. The T241A significantly affect MS, the G9457A significantly affect BF and CW, the C9662T significantly affect CW and G9936A, T9939C, G9998A significantly affect BF (P<0.05) (Table 4). And 3 polymorphisms parsed into LD block in A1M region. Previous studies have reported that several SNPs in the QTL region that influence carcass traits (CW, LMA, MS) are mapped at the region on BTA8 where AMBP gene is located (McClure et al., 2010). In pig, QTLs for meat quality traits (meat color, water content, drip loss and pH and conductivity) have been mapped in the region on SSC1 where AMBP gene in located (Liu et al., 2009). The SNP of AMBP gene was associated with meat quality traits in pig (P<0.05) (Cinar et al., 2012). And a partial protein sequence alignment of amino acid sequences from cattle, pig, human and rat, the alanine altered by T148C is conserved in all species (Fig. 3). This conservation suggests that T148C may alter economic traits in Hanwoo. These results provide supporting evidence that the AMBP gene may be a marker for carcass traits in Hanwoo.

These results suggest that the SNPs on the AMBP gene influence economic traits in Hanwoo. And it also provide evidence that the AMBP is a candidate gene for economic traits.

