
Introduction
Becasuse of centuries of domestication and breeding, a wide range of livestock breeds exists today. Duck is one of the most widely distributed livestock species in Indonesia. It plays an important role in providing income and food sources. To date, more than eight local duck breeds have been raised by livestock keepers across Indonesian archipelago. Indonesian local duck breeds seem to possess enormous genetic diversity as the result from various populations and production system developed over time by breeders to fulfil diverse needs in various environmental conditions. Genetic diversity within farm animal species refers to the extent of genetic variation within and among the breeds, strains, and lines in order to preserve the highest intraspecific variability (Lenstra et al., 2012). The availability of wide range of biodiversity allows us for implementing genetic improvement for the local breeds. It is also an important element in sustainable use of animal genetic resource (AnGR) as it allows livestock keepers to adapt their animals to changing conditions (FAO, 2007). Effective management of this AnGRs therefore, is needed to be implemented. In additon, understanding the genetic diversity among local breeds or populations, as well as the biological mechanism underlying genetic diversity can increase the successful preservation and utilization of these valuable genetic resources.
Recently, modern technique in molecular genetics has provided a number of genetic markers which can be used to asses the genetic diversity among domestic animals. The genetic markers have been comprehensively exploited to access genetic variability as they contribute information on every region of the genome, regardless of the level of gene expression (Sharma et al., 2015). Among them, microsatellite marker is the most widely used in genetic diversity studies. Microsatellites, or simple sequence repeats (SSRs), represent codominant molecular genetic markers, which are abundantly distributed across genomes (Chistiakov et al., 2006). Because of the relative ease of scoring and ability to exhibits high level of polymorphisms as well as higher heterozygosities, its application as a genetic appraisal tool is quite significant (Olowofeso et al., 2005). Microsatellite variations have been commonly used for assesing genetic diversity, differentiation, structure, and relationship among indegenous duck breeds, as well as inbreeding levels in many countries (Wu et al., 2008; Ismoyowati and Purwantini, 2011; Mukesh et al., 2011; Gaur et al., 2016; Seo et al., 2016).
To date, a few studies have reported the genetic diversity among Indonesian local duck populations (Ismoyowati and Purwantini, 2011; Rusfidra et al., 2013). The objective of this study was to evaluate the genetic diversity among Indonesian duck populations across Java island using microsatellite markers. Study on genetic diversity of Indonesian local ducks is expected to provide scientific information for designing breeding strategies and conservation plans.
Materials and Methods
Duck samples and DNA Isolation
A total of 120 individuals from 4 Indonesian local duck populations were examined (30 individuals by population). The samples were collected from different breeding regions, Mojosari (MJ) from BPTU-HPT Pelaihari, South Kalimantan; Magelang (MG) from Magelang, Central Java; Rambon (RM) from Cirebon, West Java; and Turi (TR) from Bantul, Special Region of Yogyakarta region (Figure 1). MJ population was reared under controlled breeding program, while the remaining populations were reared by traditional production system by local farmers. The blood samples were obtained from the ulnar vein. Genomic DNA was extracted from blood samples using the gSYNC DNA Extraction Kit (Geneaid, Taiwan) following the manufacturer’s instructions and stored at -20°C before doing PCR amplification.
PCR and microsatellite genotyping
Twenty two microsatellite markers were used to asses the genetic diversity (Table 1). The microsatellites were amplified with multiplex PCR using extracted genomic DNA in total volume of 20 µL. The PCR mixture contained 10 ng/μL of duck genomic DNA, 2X Multi HS Prime Taq Premix (Genetbio, Korea), 8 pmol of each fordward primers modified by four types of fluorescent dye (FAM, VIC, NED, PET) and reverse primer (Applied Biosystems, USA), and distillated water. The PCR was performed in an initial denaturation at 95°C for 10 min, followed by 38 cycles of 30 sec of denaturation at 95°C, 30 sec of annealing at 60°C, 30 sec of extension at 72°C, and final extension at 72°C for 10 min using BIO-RAD T100TM Thermal Cycler. The genotyping reaction contained of 1 μL of diluted PCR products, 10 μL of Hi-Di™ Formamide (Applied Biosystems, USA), and 0.1 μL of GeneScan™-500 LIZ™ size standard marker (Applied Biosystems, USA). Fragment analysis was performed using the Genetic Analyzer 3730 xl (Applied Biosystems, USA). The fragment size of the PCR products were identified using GeneMapper ver.3.7 (Applied Biosystems, USA).
Genetic diversity analysis
The genotyping data were used to analyze genetic diversity among populations. Number of allele (Na), expected heterozygosity (Ho), observed heterozygosity (Ho) and polymorphism information content (PIC) were analyzed using Cervus 3.0 program (Marshall et al., 1998). The F-statistics, including inbreeding coeffcient of an individual relative to the subpopulations (FIS), inbreeding coefficient of an individual relative to the total population (FIT), genetic differentiation index between population (FST) were calulated using GenAlex version 6.501 (Peakall and Smouse, 2012).
Results
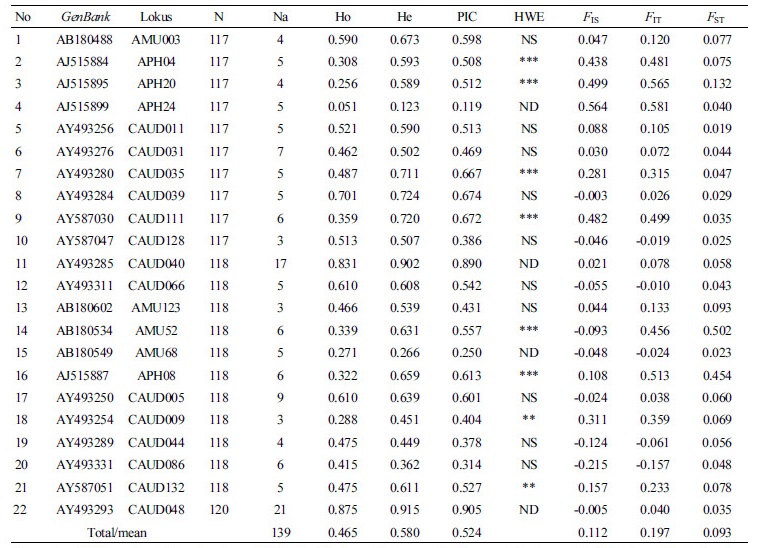
Number of alleles per locus (Na), observed heterozigosity (Ho), expected heterozigosity (He), and polymorphism information content (PIC) for each locus across all populations were summarized in Table 1. A total of 139 alleles were detected across 22 microsatellite loci, ranging from 3 (CAUD128, AMU123, CAUD009) to 21 (CAUD048), with a mean value of 6.32 alleles per locus. The Ho and He values in all loci ranged from 0.051 (APH24) to 0.875 (CAUD048) with a mean value of 0.465, and 0.123 (APH24) to 0.905 (CAUD048) with a mean value of 0.580, respectively. The PIC values for each locus varied from 0.119 (APH24) to 0.905 (CAUD048), with a mean value of 0.524. For the Wright’s F-statistics, the mean value of FIS, FIT, and FST among all loci were 0.112, 0.197, and 0.093, respectively (Table 1). All twenty two loci utilized in the present study were found to be polymorphic in all populations studied.
Discussion
To evaluate the genetic diversity among duck populations, the number of alleles, Ho, He and PIC values are the basic measures, providing important information for the discrimination of individuals and populations (Seo et al., 2016). The level of variation depicted by the number of alleles at each locus serves as a measure of genetic variability having direct impact on differentiation of breeds within a species (Buchanan et al., 1994). The mean number of alleles further reflected the medium to high level of genetic diversity among studied populations. The mean number of alleles in all loci across all analyzed populations (Na=6.95) were lower than those of Beijing duck (Na=10.7, Wu et al., 2009), Southeast Asian ducks (Na=9.38, Seo et al., 2016), and Asian ducks (Na=11.5, Sultana et al., 2017).
In our study, the mean value of observed heterozigosity (Ho) and expected heterozigosity (He) in all loci and populations also revealed medium to high genetic diversity, with the value 0.465 and 0.580, respectively. Furthermore, based on the PIC value, there were 14 microsatellite loci which had high PIC values (PIC>0.5), indicating that these microsatellite loci had high diversity and can be employed to reflect the genetic relationshiop among duck populations on molecular level. Bolstein et al. (1980) classified the PIC value as highly informative (PIC>0.50), reasonably informative (0.50>PIC>0.25), and slightly informative (PIC<0.25). Using the same microsatellite loci, the PIC values in this study (Table 1) was lower than reported by Seo et al. (2016) in Southeast Asian ducks (PIC=0.584) and Sultana et al. (2017) in Asian duck populations (PIC=0.602). Interestingly, the CAUD048 had the highest PIC value (0.905) in this study. The results were similiar to those of some studies in which PIC values of the CAUD048 were also high with value 0.932 (Seo et al., 2016) and 0.942 (Sultana et al., 2017). It suggested that CAUD048 had high diversity and discriminating power to distinguish among individuals and populations. In contrast, the APH24 had the lowest PIC value (0.119), whereas the number of alleles detected in this locus was 5. In previous study, null (0) alleles and 0.000 for PIC in the APH24 of Alabio and Bali ducks had been reported (Ismoyowati and Purwantini, 2011). Moreover, the PIC value of APH24 was 0.502 in Southeast Asian ducks (Seo et al, 2016), and 0.602 in Asian duck populations (Sultana et al., 2017), suggesting that the APH24 had lower variation in Indonesian duck populations compared to Asian duck populations.
However, the observed heterozigosity in all loci were lower than expected value, indicating that microsatellite loci pointed towards the deficit of heterozygotes and a departure from Hardy-Weinberg equilibrium (HWE). The test for HWE confirmed that in 8 out of 22 loci significantly deviated from HWE (Table 1). Various factors can contribute towards deficit of heterozygotes. First, the locus is under selection. Second, ‘null alleles’ may be present which are leading to a false observation of excess homozygotes. Third, inbreeding may be common in the population. Fourth, the presence of population substructure may lead to Wahlunds' effect (Nei, 1987; Peter et al., 2007) and genotyping errors likely due to low sample quality (Morin et al., 2009). The deviation from HWE indicated that these loci might were under selection pressure and the occurence of inbreeding in some loci across all populations.
In additon, we also evaluated the genetic diversity among populations using Wright’s F-statistic. The F-statistic (FIS, FIT, FST) give an explanation in terms of inbreeding coefficient and population differentiation. The inbreeding coeffcient of an individual relative to the subpopulations (FIS), and inbreeding coefficient of an individual relative to the total population (FIT) may be positive values or negative values, whereas, the estimated value of genetic differentiation index between population (FST) were always positive. Wu et al. (2008) reported that when there is no differentiation, the value of FST is 0; when alleles among populations are quite different, the value of FST equals 1. In this study, the overall FIS value (0.112) showed that the loci observed were higher than zero. Only 7 out of 22 loci had negative value for FIS (Table 1). The APH24 had the highest coeficient inbreeding with positive FIS value (0.659) indicating the presence of inbreeding and low heterozygotes in this locus.
Moreover, the mean FST value of 0.093 among all duck populations indicated that only 9.30% of the genetic variation was between the populations, while 90.70% was within the populations. Furthermore, based on the results suggested that there was a moderate genetic differentiation among these duck populations. The level of genetic differentiation among Indonesian duck populations was higher to those reported in Chinese (Beijing and Cherry Valley) ducks, FST = 0.075 (Wu et al., 2009) and Chinese native chicken, FST = 0.013 (Ding et al., 2010), but lower than Chinese indigenous laying-type ducks, FST = 0.184 (Su and Chen, 2009), and quite similiar to Asian duck populations, FST = 0.135 (Sultana et al., 2017).
Overall, among duck populations studied, deficit heterozygotes and low inbreeding level have been observed. The most possible reason being related to this phenomenom was that in all populations (except Mojosari) were likely in uncontrolled mating by using unequal sex ratio of breeding ducks. In farmer’s level furthermore, to improve the performance of the ducks in producing egg, they also practiced a selection based on plumage colour or production performance. The farmers selected the plumage colour as they expected the ducks which have a certain plumage colour, would produce higher eggs in quantity and quality. Therefore, this non random mating might could result on deficit heterozygotes. Mating system between the ducks was also still unclear as many of them didn’t have any pedigree record, as well as limited stock of males in the flocks, leading to the uncontrolled use of males and females in mating practices.
In conclusion, all microsatellite markers used in this study could reflect medium genetic diversity and differentiation among duck populations of Indonesia in Java island. Inbreeding coeffcient of an individual relative to the subpopulations (FIS) was positive in several duck populations. It is therefore needed to establish a well-planned and organized breeding system to avoid inbreeding depression. The gene diversity level in all populations furthermore, can provide basic information for breeding and conservation strategies to protect this prestigious gene diversity among Indonesian duck populations. However, combining the valuable genetic information from molecular analysis using these microsatellite markers and phenotypic traits analysis should be taken together as a valuable information to initiate the conservation and preservation program of genetic resources among these duck populations.

