
Introduction
Boerka, which is a crossbred goats derived from Boer buck and Kacang does has been widely raised by livestock keepers in Indonesia. Although it has 33-48% higher growth performance and greater carcass characteristics compared to Kacang goat (Ginting and Mahmilia, 2008), it is necessary to improve their production efficiency to become more competitive with other local goat breeds. Genetic improvement of local goats has been achieved using traditional breeding methods based on phenotypic information, but a wide magnitude of variability in productive performance is still present within the breed. Therefore, characterization of candidate genes associated with particular traits of interest is important to improve the accuracy of traditional breeding methods.
Meat quality traits which are controlled by many genes are among the major concern of goat profitability. Calpastatin (CAST) gene is one of the most documented gene for meat quality traits. CAST is a protein inhibitor of the ubiquitous calcium-dependent proteases, µ-calpain and m-calpain. In goat, CAST gene is located in chromosome 7 with length size of 134 Kb (14434087...14568155 based on GenBank acc NC_030814.1) and consists of 34 exonic regions. The gene encodes a specific inhibitor of the calpain, which further decreases the rate of myofibrillar protein degradation during post-mortem (Corva et al., 2007; Koohmaraie et al., 1995; Singh et al., 2012). Considerable findings demonstrate that CAST gene is a strong candidate gene for meat quality traits in goats (Khan et al., 2012; Jahromi et al., 2015; Othman et al., 2016; Bahrampour and Mohammadi, 2017). Therefore, characterization of caprine CAST gene is important for designing marker-assisted selection (MAS) for meat quality traits.
Recently, single nucleotide polymorphisms (SNPs) are desirable DNA markers for MAS because of their high abundance and codominant inheritance. These SNPs can be used to design haplotype blocks, which are sections of chromosome with high linkage disequilibrium (LD), low haplotype diversity and low recombination rate (Phillips et al., 2003). LD is useful in the determination of the actual genes that are responsible for variation of economically important traits through association mapping in livestock populations (Van Laere et al., 2003; Grisart et al., 2004). A detailed information regarding the extent of LD in genomic regions harboring quantitative trait loci (QTLs) is important for effective applications of MAS in animal breeding schemes (Dekkers, 2004). There are no literatures about CAST gene polymorphims in Boerka goat. This study was therefore undertaken to investigate SNPs of the CAST gene for determination of haplotype block pattern and linkage disequilibrium in Boerka goats.
Materials and Methods
Sample collection and DNA extraction
Blood samples were collected from 21 Boerka goats. Approximately, 3 mL of each blood sample was collected from jugular vein using EDTA vacutainer tubes (BD Bioscience, Germany). Immediately after collection, blood samples were transported to the laboratory and stored at -4°C until further treatment. Genomic DNA was extracted from whole blood using gSYNC DNA Extraction Kit (Geneaid, New Taipei City, Taiwan) according to the manufacturer’s standard procedure. The quality of extracted DNA was checked by electrophoresis in a 1% agarose gel stained with ethidium bromide mediated in a UV transilluminator.
PCR amplification
A fragment of 620 bp of caprine CAST gene was amplified using the polymerase chain reaction (PCR) technique in a SEDI G Thermal Cycler (Wealtec Corp, USA). The primers selected were CAST_F, 5’-TGGGGCCCAATGACGCCATCGATG -3’ and CAST_R, 5’-GGTGGAGCAGCACCTCTGATCACC-3’(Othman et al., 2016). The PCR reaction was made up of 2 µL of genomic DNA, 0.5 µL of each primer, 12.5 µL MyTaqTM HS Red Mix (Bioline, UK) and 9.5 µL ddH2O to make a total volume of 25 µL. The thermal cycling included an initial denaturation at 95°C for 5 min, followed by 35 cycles of 95℃ for 60 sec, 62℃ for 60 sec, 72℃ for 2 min, with a final extension at 72℃ for 10 min. The PCR products were run on 2% agarose gel (1st BASE, Singapore) to confirm the length of the amplified fragments, and 100 bp DNA-ladder (New England Biolabs, USA) was used as a molecular weight marker. The gel was visualized under UV transilluminator. The PCR products were then sequenced using ABI 3730xl genetic analyzer (Applied Biosystems, Foster City, CA, USA).
Data analysis
The sequencing results of the CAST gene were manually checked and edited using BioEdit software (Hall, 1999) and aligned using Clustal Omega (). After alignment, SNPs were inspected and used to construct haplotype block patterns (D’prime, LD, r-square (r2) and haplotype frequency) and linkage disequilibrium using Haploview software.
RESULTS
SNPs identification
A DNA fragment (620 bp) covering the exonic and intronic regions of the CAST gene was successfully amplified (Figure 1). After alignment of 21 sequences, five SNPs were found in non-coding region (intron 12), namely g.146C>A, g.224A>G, g.281G>A, g.737C>T and g.431G>A. No polymorphisms were detected in the exonic regions. Figure 1 shows clear peaks in the electrophoregram for each detected SNP.
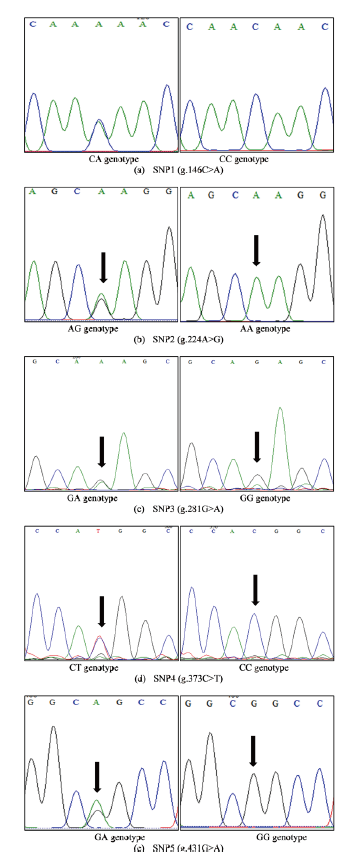
LD and haplotype block analysis
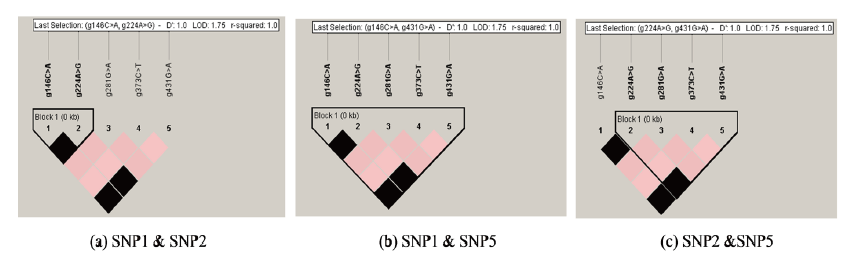
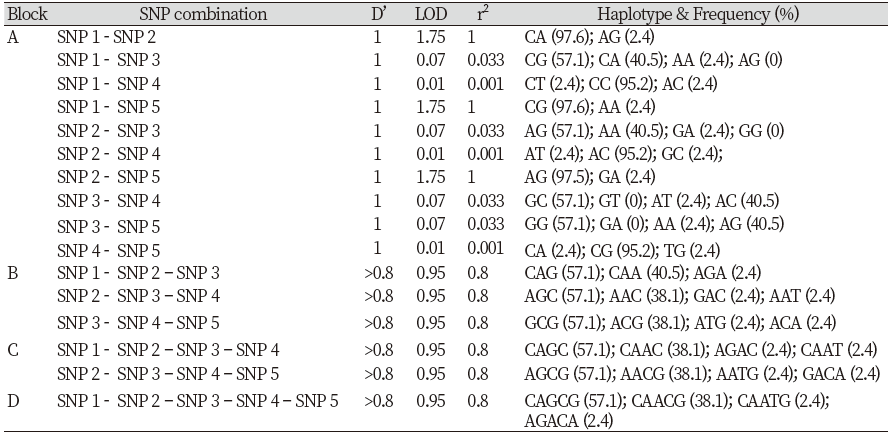
Using Haploview software the SNPs identified in this study were analyzed for LD and haplotype block patterns. Four haplotype block patterns with different pairs of loci were performed (Figure 2). All blocks had D’prime values equal to 1.0. Haplotype block A, B, C and D consisted of 10, 3, 2 and 1 SNPs combination (Table 1). The highest LOD (1.75), r2 (1.00) and haplotype frequencies (97.6%) were observed in block A (SNP1&2, SNP1&5 and SNP2&5). The lowest LD (0.01) and r2 (0.0010) were found in block A (SNP1&4, SNP2&4 and SNP4&5). Block B, C and D had only 3, 2 and 1 SNPs combination, respectively, with a medium haplotype frequency of 57.1% each. Seven and four pairs of SNPs with high (>95%) and medium (<57.1%) frequencies were observed in Block A. All pairs of SNPs in Block B, C and D were in medium frequency (57.1%).
Discussion
A 620 bp of the caprine CAST gene amplified in this study was found to be polymorphic. Sharma et al. (2013) found nine novel SNPs in intron 5, 7 and 8, while Zhou and Hickford (2008) found a missense mutation in exon 6, which leads to change amino acid from Ser to Arg in the L domain of the protein. In contrast to the results of this study, no polymorphism was observed in intron 12 of the CAST gene in Indian goat breeds (Sharma et al., 2013).
CAST gene polymorphisms in domestic animals have been studied by some researchers. Othman et al. (2016) detected polymorphisms of the CAST gene in Egyptian sheep and goats, and found two genotypes (GG and AG) in 140 animals investigated. In both sheep and goat populations, the frequency of GG genotype was higher than AG genotype (65.9% vs 34.1% and 56.9% vs 43.1%, respectively). Using PCR-SSCP technique Zhou and Hickford (2008) detected polymorphisms of the CAST gene in New Zealand Boer goat, which lead to amino acid substitution (Ser/Arg) in exon 6 in domain L of the protein. Moreover, Khan et al. (2012) observed 2 alleles, M and N, of the CAST/MspI locus in Balkhi and Kajli sheep breeds, with frequencies of MM, MN and NN genotypes being 76, 24 and 0% in Balkhi breed and 74, 24 and 2% in Kajli breed, respectively. They demonstrated that A>G polymorphism in the exon 1C/1D region of the ovine CAST gene is a potential molecular marker for MAS associated with growth rate.
Linkage disequilibrium (LD) has been widely used in localizing candidate genes associated with a particular disease in human (Van Eerdewegh et al. 2002; Rioux et al. 2001; Geesaman et al. 2003; Stoll et al. 2004). LD and haplotype block patterns can be performed using Haploview software by selecting one of several block definitions to indicate the region into segments of strong LD (Gabriel et al. 2002; Wang et al. 2002). The software allows user to select groups of markers for subsequent haplotype analysis with different color schemes to represent the LD relationship. Dark grey indicates a strong LD; light grey shows uniformative; and white color represents strong recombination. For LOD, there are four color schemes (white: LOD<2 and D’<1; shade of pink/red: LOD#2 and D’<1; blue: LOD<2 and D’=1; bright red: LOD #2 and D’=1). For r2, there are three color schemes, namely white (r2=0), shade of grey (0<r2<10) and black (r2=1). A graphical representation can be created to calculate several pairwise measures of LD and select one of commonly used block definitions.
The highest LOD (1.75), r2 (1.00) and haplotype frequencies (97.6%) were observed in block A (SNP1&2, SNP1&5 and SNP2&5). The lowest LD (0.01) and r2 (0.0010) were found in block A (SNP1&4, SNP2&4 and SNP4&5). Block B, C and D had only 3, 2 and 1 SNPs combination, respectively, with a medium haplotype frequency of 57.1% each. The LOD score is a statistical test for the analysis of linkage in human, animal and plant populations. Positive LOD score indicates the presence of linkage. Furthermore, a LOD score greater than 3 indicates evidence of linkage (Morton, 1955). All of the LOD scores in this study were positive indicating the linkage among the SNPs. Linkage disequilibrium represents a nonrandom association between pairs of alleles. The resulted LD structure can be used to determine the evolutionary history of populations.
Conclusion
Four haplotype block patterns with different pairs of loci were performed. All blocks had D’prime values equal to 1.0. Haplotype block A, B, C and D consisted of 10, 3, 2 and 1 SNPs combination. The SNPs and their haplotype combinations in the CAST gene may be useful for future association studies with meat quality traits in goats.

