Soon Hwangbo1![]() , Yangmo Koo2*
, Yangmo Koo2*![]() , Ji Hong Lee3*
, Ji Hong Lee3*![]()

The inbreeding coefficient (F) of each animal was estimated based on the path coefficient method originally proposed by Wright (1922). For efficient processing of large-scale pedigree data, the algorithm developed by Meuwissen and Luo (1992) was applied. Inbreeding coefficients were obtained from the diagonal elements of the numerator relationship matrix (A-matrix). The basic definition of the inbreeding coefficient (Fx) follows the standard formula described in previous studies.
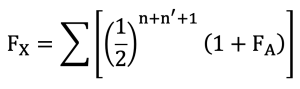
In this context, Fx represents the inbreeding coefficient of individual X, and FA represents the inbreeding coefficient of the common ancestor A. The term n indicates the number of generations from the sire to the common ancestor, while n* indicates the number of generations from the dam to the common ancestor.
The inbreeding coefficient of each individual was calculated based on the genetic relationships among all animals included in the pedigree. Estimation of inbreeding coefficients was performed using the PROC INBREED procedure of the SAS (Statistical Analysis System) package. This procedure estimates covariance between individuals by using information from all known ancestors in the pedigree. The inbreeding coefficient (Fx) of a given individual X is equal to the coancestry coefficient between its parents (S,D). This value is defined as the diagonal element of the numerator relationship matrix for individual X minus one.
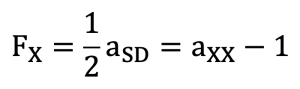
In this definition, aSD represents the additive genetic relationship between the sire (S) and the dam (D), and axx represents the additive genetic relationship between individual X and itself, corresponding to the diagonal element of the relationship matrix.
To quantify the availability of pedigree information, the pedigree completeness index (PCI) was calculated for each individual. PCI reflects whether ancestor information is recorded at each generation for a given animal. The index was evaluated by expanding pedigree depth from the first generation (sire and dam) to the second, third, and higher generations. PCI at generation t was defined as follows.
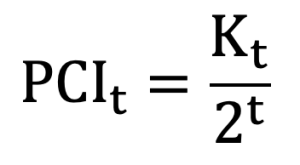
In this definition, Kt represents the number of ancestors that were actually identified at generation t, and 2t represents the total number of ancestors that are theoretically expected at that generation (Boichard et al., 1997; MacCluer et al., 1983). The PCI value for each individual was calculated as the average of PCI values across generations. This approach was used to quantify the reliability of pedigree information for individual animals. In this study, the distribution of individual PCI values was compared across birth periods based on birth year.
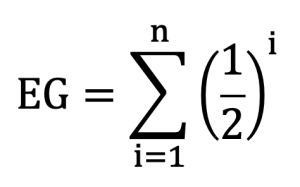
Here, i indicates the generation where parental information was still available for an ancestor. PCI values and related generation indices were used as indicators to verify the completeness of pedigree information. When interpreting the inbreeding coefficient, it was used as a criterion for determining reliability.
In order to determine the cause of the increase in the inbreeding, the contribution of common ancestors related to the pedigree was analyzed. In addition, founder genome equivalents and the actual genetic contribution of sires were analyzed. Founders were treated as animals without parental records. Founder contribution was calculated as the genetic contribution rate of individual descendant.
The contribution of each founder f was estimated according to the method described by Boichard et al.(1997).
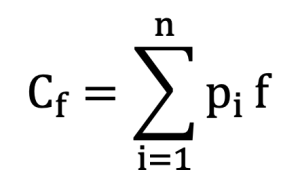
In this equation, pif represents the probability of genetic contribution from founder f to descendant i, and n represents the total number of descendant animals.
Founder genome equivalent (Fge) was calculated using the method proposed by Leroi et al.(2012).
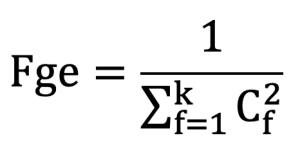
In this definition, k represents the number of founders, ![]() and represents the sum of squared genetic contributions of individual founders.
and represents the sum of squared genetic contributions of individual founders.
To examine whether changes in inbreeding in Hanwoo cow population were associated with shifts in mating patterns, several population structure indicators were evaluated. These included generation structure, frequency of sires used for mating, distribution of offspring numbers per sire, sex ratio composition, and generation interval (GI) (Dunlop et al., 1993). Generation interval was estimated based on the relationship between parents and offspring. Birth dates of sires and dams were calculated as the average birth date of their offspring, and GI was estimated accordingly.
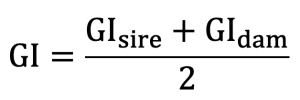
To further assess repeated use of the same sire, the average number of offspring per sire by year was calculated. In addition, same sire usage bias was evaluated using the proportion of offspring produced by the top 10% of sires.